シロイヌナズナ における, 細胞外領域にロイシンリッチリピート(Leucine- Rich Repeat,LRR)を有するLRR型受容体キナーゼ は 200種類以上 も知られているが, 大半の受容体のリガンド や機能および活性化機構 は未知のまま残されている.私たちは,LRR型受容体であるBAM1(CLV1ホモログ)のリガンドのひとつがCLE9ペプチドであることを突き止めた.さらに光反応基および放射性標識を導入したCLE9ペプチド誘導体を合成し,BAM1に対してフォトアフィニティーラベルを行なった結果,BAM1におけるリガンド結合領域がLRRの6−8番目に存在することを見出した.BAM1の詳細な配列解析および立体構造のモデリング解析から,結合領域のアミノ酸配列はLRRのコンセンサス配列から若干逸脱しており,その乱れはすべてらせん構造の内側を向いていることがわかった.リガンドはおそらくらせん構造の内側部分で認識されており,このわずかな配列の乱れが受容体のリガンド結合特異性を決定づけているものと考えられる.一部の受容体では,膜貫通領域に近いアイランド領域がリガンド結合に関わっていることが知られていたが,BAM1では,膜貫通領域から遠い位置にあるLRRにリガンドが直接結合することが示されたことで,LRR型受容体キナーゼの多様なリガンド認識機構が明らかになった.
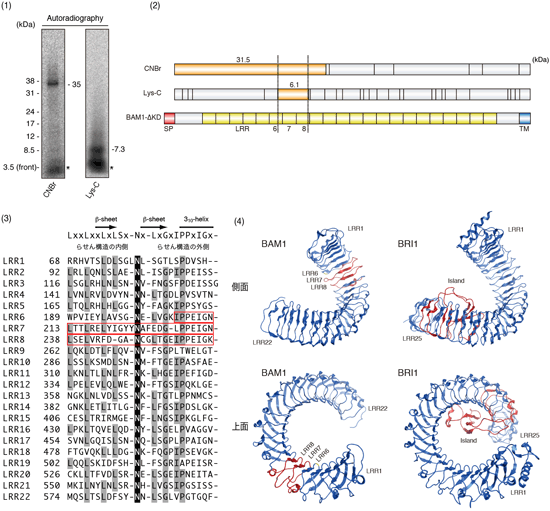
図1.BAM1のリガンド結合部位の同定
(1)ラベル導入したBAM1を断片化し,オートラジオグラフィーで検出した.(2)得られた断片の大きさと,理論上の断片の大きさを比較することで,リガンド結合部位がLRRの6–8番目に存在することが示された.(3)BAM1細胞外領域におけるLRRのアミノ酸配列.リガンド結合部位は赤線で囲われた部分.リガンド結合部位ではLRRに高度に保存されているアミノ酸の置換や欠失などがみられ,構造に若干のゆらぎが生じていると推測される.(4)BAM1立体構造のコンピュータ予測結果(左側).ブラシノステロイド受容体BRI1の立体構造(右側)と比較して,リガンド結合部位は比較的N末端に近い位置に存在することが示された.双方のリガンド結合部位は赤色で示してある. |