植物の根系は,発芽後に伸びる主根,地上部から発生する不定根,そして,これらの根から発生する側根から成り立っています.シロイヌナズナの側根は根の内鞘細胞の非対称な分裂によって形成が開始されます.これまでに私たちは,オーキシン応答で働く「SLR/IAA14-ARF7-ARF19オーキシンシグナルモジュール」を介した遺伝子発現制御が側根の形成開始に必要であり、その下流で誘導されるLBD16/ASL18を中心としたLBD/ASLタンパク質群が関わる可能性を示してきました.しかし,LBD16/ASL18タンパク質が側根の形成過程のどの時期に、どの細胞でどのような発生段階に働くのか不明のままでした.
今回,LBD16/ASL18タンパク質が根の内鞘細胞のうち,側根を生み出す細胞(側根創始細胞)において,側根形成が開始される時期に特異的に発現することを示しました.さらに,LBD16/ASL18とその類似タンパク質群の機能を同時に抑制したところ,側根創始細胞において核の移動をともなう非対称性が確立されずに等分裂が1回起こるだけで,その後の側根形成が完全に阻害されました.これらの結果から,LBD16/ASL18タンパク質や類似のLBD/ASLタンパク質群は側根創始細胞の非対称性の確立を制御することで,側根の発生を誘導していることが明らかになりました.
図1.LBD16/ASL18および類似のLBD/ASLタンパク質群の働きを抑制したシロイヌナズナ (14日目の芽生え).野生型では側根が形成されるのに対して,LBD16/ASL18の機能抑制植物では側根形成が起こらない.スケールバー:10 mm
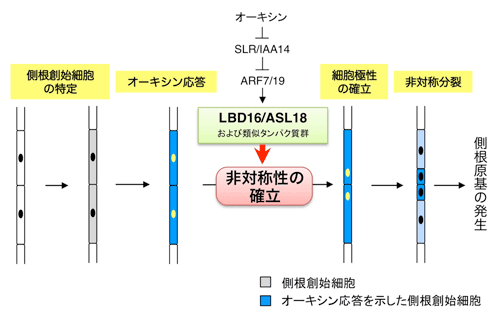
図2.側根形成開始におけるLBD16/ASL18および類似のLBD/ASLタンパク質の役割
LBD16/ASL18は側根創始細胞においてSLR/IAA14-ARF7-ARF19オーキシンシグナリングモジュールに依存して活性化され,類似のLBD/ASLタンパク質とともに,側根創始細胞の非対称性の確立を促進する.その結果,細胞核の移動と非対称分裂が起き,側根形成が開始する. |