Shigeyuki Betsuyaku, Fuminori Takahashi, Atsuko Kinoshita, Hiroki Miwa, Kazuo Shinozaki, Hiroo Fukuda and Shinichiro Sawa
(別役重之、高橋史憲、木下温子、三輪大樹、篠崎一雄、福田裕穂、澤進一郎)
論文題目
Mitogen-Activated Protein Kinase regulated by the CLAVATA receptors contributes to the shoot apical meristem homeostasis.
(CLAVATA受容体によって制御されるMAPKが茎頂分裂組織の恒常性維持に寄与する。)
発表誌
Plant Cell Physiol, 2011 vol. 52 (1) pp. 14-29
要 旨
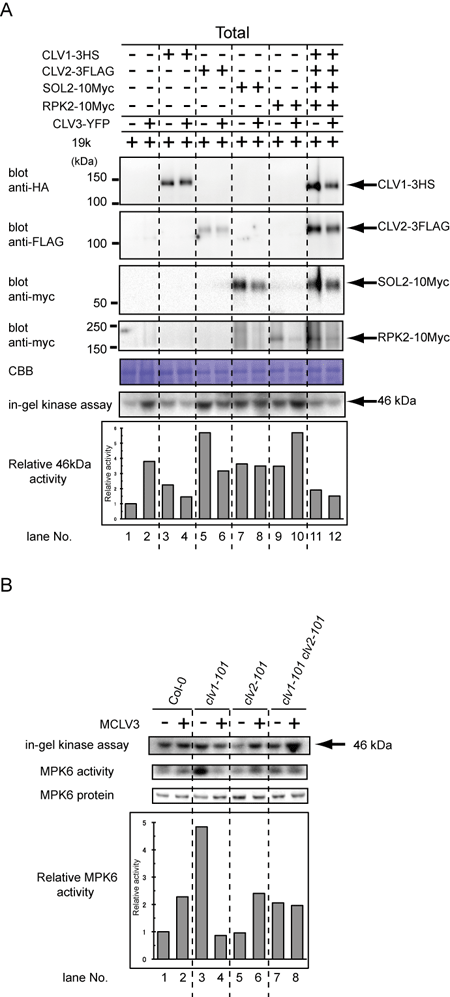
アラビドプシスの茎長分裂組織(SAM)においては、CLAVATA(CLV)経路がSAM幹細胞集団のサイズ制御を担っている。そこでは、分泌型ペプチドリガンドとして機能するCLV3がCLV1、 CLV2-SUPPRESSOR OF LLP1-2 (SOL2)/CORYNE (CRN)、RECEPTOR-LIKE PROTEIN KINASE 2 (RPK2)/TOADSTOOL 2 (TOAD2)からなる3つの受容体キナーゼ複合体を介して転写因子WUSCHELの発現を負に制御している事が遺伝学的に知られていたが、その詳細な分子機構については未知であった。