| 研究代表者 |
杉山宗隆、出村拓 |
所 属 |
東京大学大学院理学系研究科附属植物園 |
| 著 者 |
Ohtani, M., Demura, T., Sugiyama, M.
(大谷美沙都、出村拓、杉山宗隆) |
| 論文題目 |
Particular significance of SRD2
-dependent snRNA accumulation in polarized pattern generation
during lateral root development of Arabidopsis.
(シロイヌナズナの側根発生時の極性パターン形成においてSRD2に依存したsnRNA蓄積がもつ特別な意味) |
| 発表誌 |
Plant & Cell Physiology in press |
| 要 旨 |
側根の形成は、内鞘細胞の垂層分裂から始まる。一連の秩序だった細胞分裂によって側根の原基が完成すると、細胞分裂が停止し、その後根端メリステムの活性化が起きる。私たちはシロイヌナズナを用いてこの過程を調べ、核内低分子RNA(snRNA)転写活性化因子をコードするSRD2
遺伝子の温度感受性変異srd2-1 が細胞分裂の制御に強く影響することを見出した。srd2-1
変異体を制限温度下で培養すると、側根原基における細胞分裂パターンが乱れるとともに、原基的な分裂が長期に亘って持続し、側根が瘤化した。このとき、基部-先端方向、放射方向いずれの軸も確立していなかった。また、srd2-1
変異体の側根原基では、オーキシン排出に関わるPINタンパク質の発現が減少しており、おそらくはそのためにオーキシン濃度勾配の形成が妨げられていた。スプライソソームsnRNAは、側根原基全体に大量に存在したが、細胞分裂の停止に際して著しく減少し、メリステムが活性化するときに再び増大した。srd2-1
変異はこうしたsnRNAの蓄積を強く阻害した。以上の結果より、側根形成過程での極性軸確立に関与するPINの発現に、SRD2
の機能に依存した高レベルのsnRNA蓄積がきわめて重要であることが示唆された。
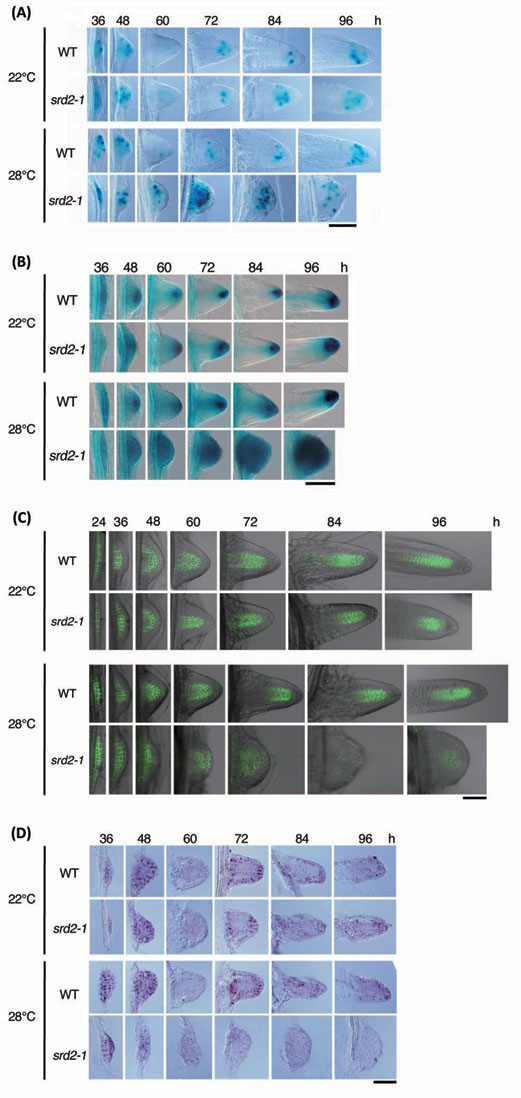
図 側根形成過程での細胞分裂、オーキシン分布、PIN1発現およびsnRNA蓄積に対するsrd2
変異の影響
野生型(WT)とsrd2-1 変異体の芽生えから茎頂部と根端部を取り除き、インドール酪酸を含む培地に置床して22°Cまたは28°Cで培養を行い、半同調的な側根形成を誘導した。
(A)CYCB1;1::DB:GUS をマーカーとして側根形成過程での細胞分裂パターンを調べた。青く染まっているのが分裂細胞。正常な側根形成の場合、活発な細胞分裂によって原基が発達した後、細胞分裂が停止し、根端メリステムでの先端成長に切り替わるが、高温条件で培養したsrd2-1
変異体では、側根原基全域での細胞分裂が長く続いた。
(B)DR5::GUS をマーカーに用い、オーキシンの分布を調べた。青い染色が濃いほど、オーキシン濃度が高い。正常な側根形成過程では、側根原基の発達につれて、先端にピークをもつオーキシン濃度勾配が形成される。これに対し、高温条件で培養したsrd2-1
変異体では、原基全体で一様にオーキシン濃度が高く、明瞭な濃度勾配は認められなかった。
(C)PIN1::PIN1:GFP をレポーターとして、PIN1タンパク質の発現と分布を調べた。緑色の蛍光がPIN1タンパク質の存在を示す。高温条件で培養したsrd2-1
変異体ではPIN1タンパク質が著しく減少しており、これがオーキシン濃度勾配の不成立と関わっていることが窺われた。
(D)スプライソソームsnRNAの一種U2 snRNAの蓄積パターンを、in situハイブリダイゼーション法により調べた。紫色のシグナルがU2
snRNAの蓄積を示す。正常な側根形成の場合、発達中の原基では全域に大量のsnRNAの蓄積が見られたが、原基の構造が完成して細胞分裂が停止する時期になるとsnRNAは大きく減少し、メリステムが活動を始める時期に再び増大した。高温条件で培養したsrd2-1
変異体では、ごく初期の側根原基を除くと、snRNAの蓄積のレベルは一貫して非常に低かった。これはSRD2
遺伝子の機能低下の直接的影響と考えられる。
スケールバーはすべて50 μm。
|
| 研究室HP |
http://www.bg.s.u-tokyo.ac.jp/koishikawa/research/sugi-lab/sugi-1.html
|