論文紹介
| 研究代表者 |
山本興太朗 |
所 属 |
北海道大学・大学院理学研究院・生命理学部門 |
| 著 者 |
S. Tashiro, C.-e. Tian, M. K.
Watahiki, K. T. Yamamoto
(田代智子、田長恩、綿引雅昭、山本興太朗) |
| 論文題目 |
Changes in growth kinetics of
stamen filaments cause inefficient pollination in massugu2,
an auxin insensitive, dominant mutant of Arabidopsis thaliana.
(シロイヌナズナ雄蘂花糸の成長速度はMSG2のオーキシン非感受性優性変異によって変化し、それが同変異体の受粉効率低下をもたらす
) |
| 発表誌 |
Physiologia Plantarum,
137, 175-187, 2009. |
| 要 旨 |
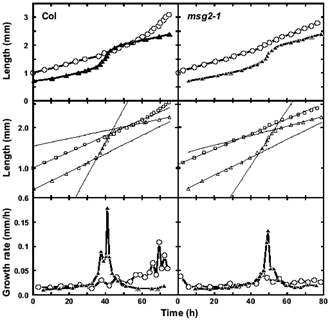
オーキシン一次応答遺伝子MSG2/IAA19の優性突然変異体msg2の稔性は野生型より低下しているので、その原因を調べた。シロイヌナズナの花では、開花頃になって花糸(雄蘂の軸にあたる部分)の成長速度が急激に上昇して雄蘂が雌蘂を追い越すようになり、その時、柱頭と葯が接触して受粉が起こる。この成長速度の上昇がmsg2ではそれほど起こらず、起こっても野生型より遅いタイミングで起こるので、雄蘂が雌蘂を追い越すことができず、その結果、受粉が起こりにくい(図1)。この研究をおこなって一番おもしろかった点は、元気なシロイヌナズナでは花の形成から開花、落花に至る過程が定常状態になっているため、ある時点で多数の花の雌蘂と雄蘂の長さの関係を測定すると、それだけでその二つの器官の相対的な成長速度が計算できると分かったことである。研究を始める前にはこのことに気がついていなかったが、データを眺めている内にこの単純な事実に気がついた。論文の考察を書くときにSmyth
et al. (1990)を読み直してみたら、彼らは既にこのことに気がついていて(花を観察していたら気がつくのは当然であるが)、花が各発達段階を経過するのに必要な時間を推定するのに利用していた。この他、この成長速度変化にMSG2の発現レベルが対応していること(図2)、花糸伸長に働いているオーキシン応答因子はARF6と8であること、花糸の表皮細胞は成長の間、細胞分裂をしないことを発見した。
|
|
|
図1 野生型(左)とmsg2突然変異体(右)の花の雌蘂(○)と雄蘂花糸(△)の成長曲線(上、中)と成長速度(下)。成長曲線は長さを線型(上)と対数(中)スケールで表示。雌蘂の長さが1.0
mmに達した時点以後の変化を調べた。msg2では最大成長速度が低下していることと、成長速度が増加するタイミングが遅れていることがわかる。
|
|
|
|
図2 MSG2プロモーターGUSシロイヌナズナの花序のGUS染色像。ある程度長く伸長した花糸だけに強いシグナルが見られる。若い花芽の葯にも弱いシグナルがあることがある。AとBのスケールバーは各々2と1mm。Aの番号は花の発達段階を表す。
|
|
| 研究室HP |
http://www.sci.hokudai.ac.jp/~ky082/keitai1/kty/index.htm |
|