論文紹介
| 研究代表者 |
荒木 崇 |
所 属 |
京都大学 大学院生命科学研究科統合生命科学専攻 |
| 著 者 |
Ikeda, Y., Kobayashi, Y., Yamaguchi, A., Abe, M., Araki, T.
(池田陽子、小林恭士、山口礼子、阿部光知、荒木 崇) |
| 論文題目 |
Molecular basis of late-flowering phenotype caused by dominant epi-alleles of the FWA locus in Arabidopsis.
(シロイヌナズナFWA遺伝子の優性エピアレルによる花成遅延表現型の分子基盤) |
| 発表誌 |
Plant & Cell Physiology 48, 205-220 (2007). [erratum: ibid 48, 562 (2007)] |
| 要 旨 |
シロイヌナズナの優性の花成遅延変異fwaは、FWA遺伝子のプロモータ領域の低メチル化により、HD-ZIP蛋白質FWAが異所発現することによるものであることが知られている。しかし、HD-ZIP蛋白質FWAの異所発現がなぜ花成の遅延を引き起こすのかは不明であった。また、そもそもFWA遺伝子は花成の制御において何らかの役割をになっているのかも明らかではなかった。本研究では、まず、FWA遺伝子は栄養成長期の植物では全く発現しないことを示し、FWA遺伝子が花成制御において何らかの役割を果たしている可能性を否定した。さらに、FWA蛋白質が花成の重要な促進因子であるFT蛋白質と特異的に結合することを示し、蛋白質間相互作用を介したFT蛋白質の機能阻害が花成遅延の主要な原因であることを示した。これを利用して、組織特異的なFWA蛋白質の異所発現により、植物体内でFT蛋白質の機能が必要とされる組織は茎頂であることを示した。FWA蛋白質と同じサブファミリーのHD-ZIP蛋白質は転写調節因子として機能することが示されていることから、FWA蛋白質の異所発現によって遺伝子発現制御の撹乱が起こることが予想された。しかし、fwa変異体と野生型の間で、遺伝子発現プロファイルの変化はみとめられず、FWA蛋白質の転写調節因子としての機能は失われていることが示唆された。
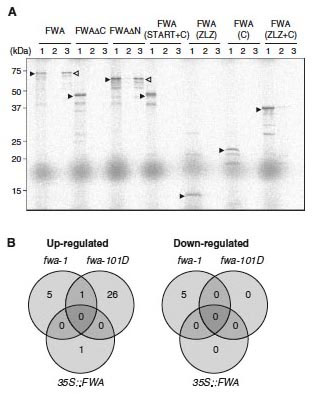 図1(A)
FWA蛋白質の全長およびさまざまな部分欠失型の蛋白質とFT蛋白質の結合。FT蛋白質としてFT-GST融合蛋白質を用い、GST-pull
downアッセイにより、放射標識した各種FWA蛋白質の結合を調べた。レーン1は標識蛋白質(レーン2, 3の1/10量)、レーン2はGSTによるpull
down、レーン3はFT-GST融合蛋白質によるpull down。 全長蛋白質(FWA)およびホメオドメインを含むN末領域のの欠失体(FWAΔN)はFT蛋白質と結合する。
(B) FWA遺伝子を異所発現する3つの系統における遺伝子発現プロファイルの変化。2つの独立の優性fwa変異体(fwa-1, fwa-101D)および、過剰発現形質転換体(35S::FWA)のそれぞれについて、野生型に対して発現量が4倍以上(Up-regulated)あるいは1/4以下(Down-regulated)になっている遺伝子の数を示す。FWA遺伝子を異所発現する複数の系統で共通して発現が変化している遺伝子がほとんどないことがわかった。 |
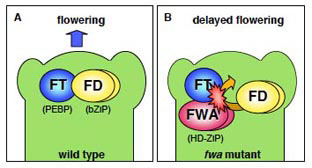 図2 (A)
野生型植物では、FWA蛋白質は発現せず、FT蛋白質(フロリゲン)とbZIP転写因子の蛋白質間相互作用により花成が促進される。
(B) 優性のfwa変異体では、FWA遺伝子のプロモーター領域の低メチル化により発現したFWA蛋白質がFT蛋白質と結合することで、茎頂におけるFT蛋白質の機能を阻害する。これにより、花成の遅れがおこる。 |
|
| 研究室HP |
http://www.lif.kyoto-u.ac.jp/labs/plantdevbio/ |
|