論文紹介
| 研究代表者 |
杉山宗隆、田坂昌生 |
所 属 |
東京大学大学院理学系研究科
附属植物園 |
| 著 者 |
Tamaki, H., Konishi, M., Daimon, Y., Aida, M., Tasaka, M., Sugiyama, M.
(玉置裕章、小西美稲子、大門靖史、相田光宏、田坂昌生、杉山宗隆) |
| 論文題目 |
Identification of novel meristem factors involved in shoot regeneration through the analysis of temperature-sensitive mutants of Arabidopsis.
(シロイヌナズナの温度感受性変異体の解析を通して同定したシュート再生に関与する新規メリステム因子) |
| 発表誌 |
The Plant Journal 57: 1027-1039 (2009) |
| 要 旨 |
植物の組織培養による器官再生は頂端メリステムが新たに形づくられる過程であり、メリステムの遺伝子ネットワークに関して重要な情報を提供する。私たちはシロイヌナズナの温度感受性変異体、rid3とrgd3を用いてシュート再生の解析を行い、シュート頂メリステム(SAM)の新形成に関わる新規因子を同定した。カルスからシュート再生を誘導すると、すぐに細胞分裂が停止したが、その後カルス表面で局所的に分裂が再活性化して細胞集塊が生じ、やがてそこにSAMが構築された。rgd3変異はこの細胞分裂再活性化を阻害し、SAM制御遺伝子のCUC1、CUC2、STMの発現を抑制した。一方、rid3変異はカルス表層で過剰な細胞分裂とCUC1発現の昂進・拡大を引き起こした。ポジショナルクローニングにより、RGD3はTATA結合タンパク質付随因子の一種であるBTAF1、またRID3は未解析のWD-40リピートタンパク質をコードしていることが判明した。シュート再生の初期段階において、RGD3はCUC1とともに発達中の細胞塊に発現し、RID3は細胞塊以外の領域に発現していた。また、RID3の強制発現は、CUC1とSTMの発現レベルを著しく低下させた。これらの結果から、SAM新形成の基盤となる細胞分裂の適切な調節に、CUC-STM経路のRID3による抑制的制御とRGD3による促進的制御が関与することが示された。
|
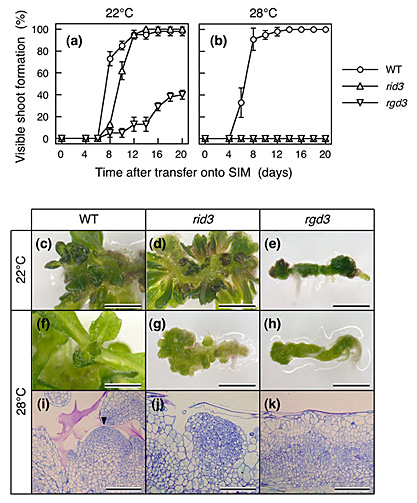
図1 rid3とrgd3における温度依存的なシュート再生不全
野生型(WT)と変異体(rid3およびrgd3)の胚軸外植片をカルス誘導培地(CIM)に置床し22°Cで前培養を行った。その後に外植片をシュート誘導培地(SIM)に移植して22°Cまたは28°Cで培養し、シュート再生を誘導した。
(a,b)22°C(a)および28°C(b)におけるシュート再生率の時間経過。rid3、rgd3のどちらにおいても、シュート再生は明瞭な温度感受性を示し、高温条件でシュート再生が強く阻害された。
(c-h)SIMに移植後、22°C(c-e)または28°C(f-h)で14日間培養した野生型(c,f)、rid3(d,g)、rgd3(e,h)の外植片の外観。28°Cで培養したrid3の外植片では多数の瘤が見られた。
(i-k)SIMに移植後、28°Cで10日間培養した野生型(c,f)、rid3(d,g)、rgd3(e,h)の外植片の組織学的観察。rid3のカルス表面では増殖細胞の集塊が異常に大きく発達していたが、rgd3のカルス表層の細胞は一様に液胞化していた。 |
|
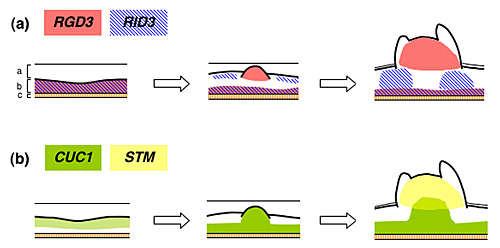
図2 シュート再生過程におけるRID3、RGD3、CUC1、STMの発現パターン
左端はSIMに移植した直後の胚軸外植片。aは胚軸の表皮と皮層、bは中心柱に由来するカルス、cは維管束を表す。この段階ではRID3とRGD3はカルス全体に強く発現しており、CUC1の発現はカルス表層部で高まっている。SIMで数日間培養すると、中央の図に示したように、カルス表層におけるRGD3とCUC1の発現は発達中の細胞塊に限局化してくる。このときRID3は細胞塊を避けるような発現パターンをとる。右端に示したように、最終的には細胞塊からSAMが構築されるが、このSAMの部分ではRGD3とSTMの発現が顕著である。 |
|
| 研究室HP |
http://www.bg.s.u-tokyo.ac.jp/ |
|