|
脳回路構造学 Group of Neural Circuit 上川内研究室 |
|
|---|
|
名古屋大学 大学院理学研究科 生命理学領域 脳回路構造学 〒464-8602 名古屋市千種区不老町 |
研究内容について私たちは、「ショウジョウバエ」 と 「蚊」 を使って、求愛コミュニケーションを成立させる脳のしくみを調べています。最近の研究発表■ プロポーズの流儀を決める遺伝子と脳のスイッチTanaka*† , Hara †, Sato, Kohatsu, Murakami, Higuchi, Awasaki, Kondo, Toyoda, Kamikouchi , Yamamoto* (2025).  ■ メスの求愛歌受け入れをドーパミンが促進する Yamakoshi, Horigome, Yamamoto, Iwanami, Iwami, Tanaka, Ishikawa, & Kamikouchi (2025)  ■ 蚊の"聴覚"を操る鍵は、神経伝達物質と細胞内シグナル Xu, Loh, Lee, Chen, Loh, Ohashi, Eberl, Andrés, Su#, Kamikouchi# (2025)  ■ 蚊の脳が音に反応する様子を世界で初めて可視化 Ohashi, Xu, Shigaki, Nakamura, Lee, Loh, Mishiro-Sato, Eberl, Su* & Kamikouchi* (2025)  ■ 蚊の異種交配を抑制する聴覚特性の種間差 Loh#, Xu#, Lee, Ohashi, Zhang, Eberl, Su* & Kamikouchi* (2024)  ■ ハエのリズム聴き分け能力を上げる神経機構 Imoto, Ishikawa, Aso, Funke, Tanaka & Kamikouchi (2024)  ■ 2種類のハエの求愛歌の好みの進化を支える神経機構 Ohashi, Ishikawa, Awasaki, Su, Yoneyama, Morimoto & Kamikouchi (2023) 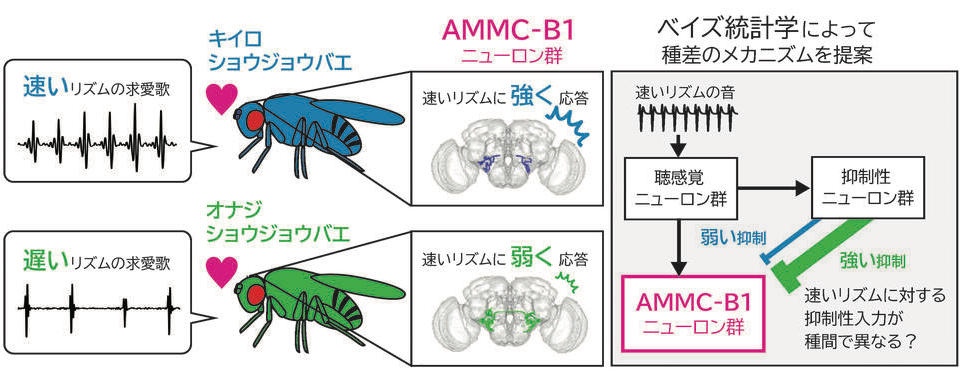 ■ 交尾を継続するためのメカニズム Yamanouchi, Tanaka & Kamikouchi (2023) 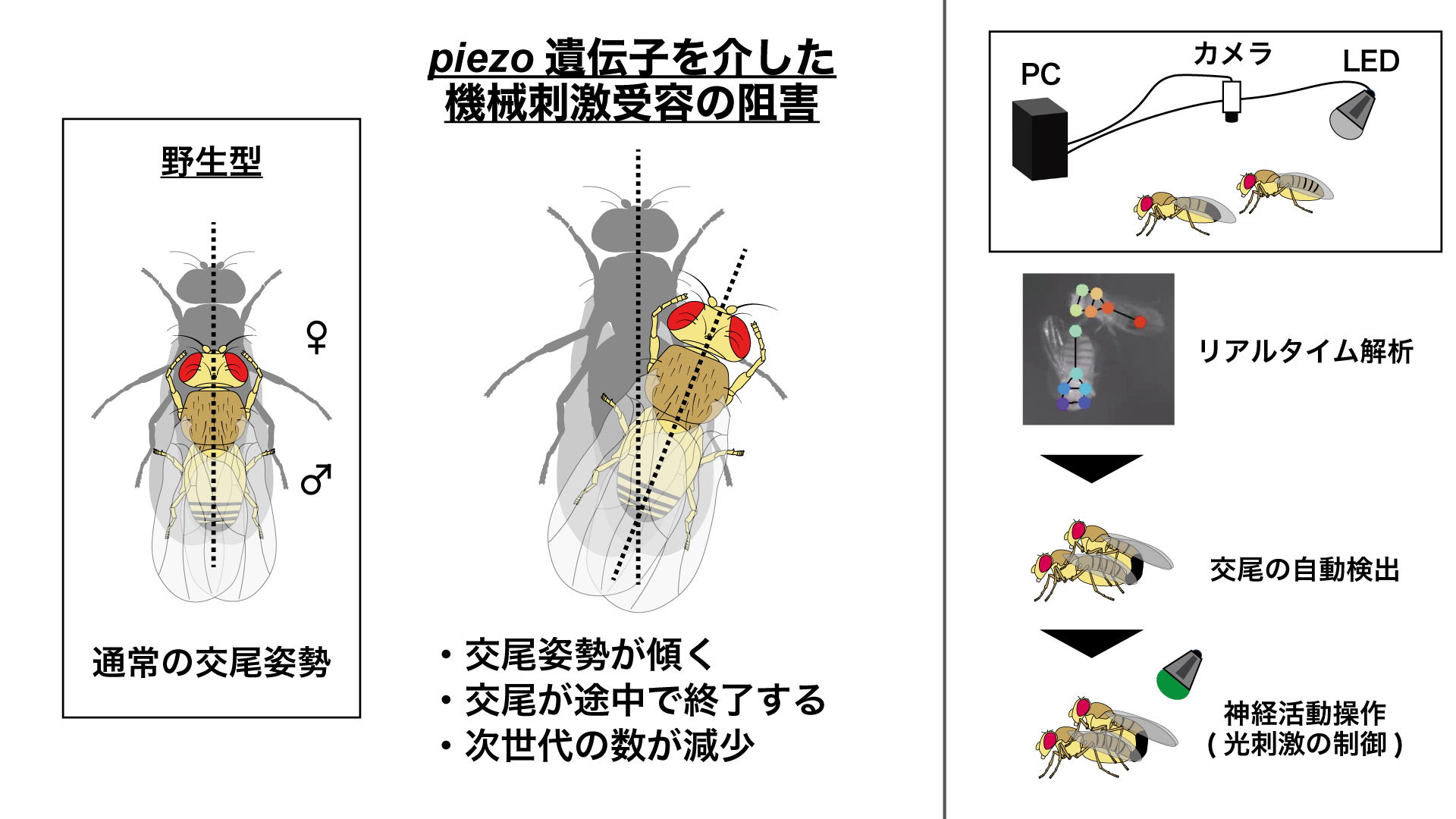 ■ 蚊の「耳」を操る物質を発見 Xu, Loh et al., (2022) 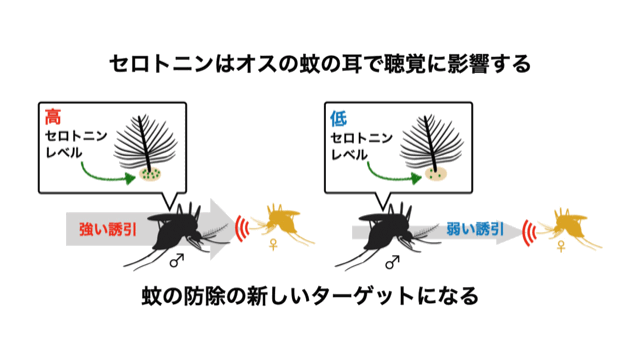 ■ 2種類のハエが示す、群れ形成のダイナミクスの違い Shirasaki, Tanaka et al., (2022)  ■ 音のリズムを聞き分けるメカニズムの発見 Yamada et al., (2018) 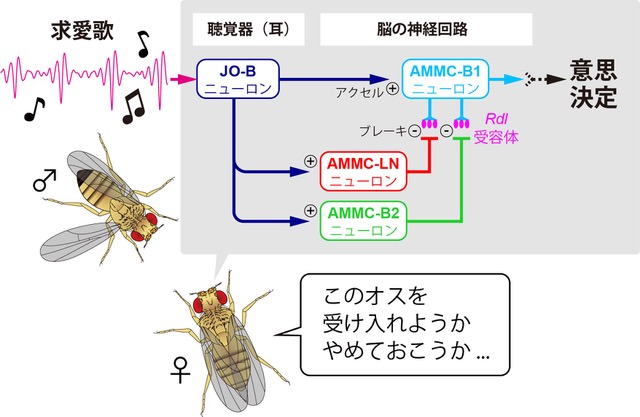
■ ハエの歌識別学習の発見 Li et al., (2018) 
■ ショウジョウバエ聴覚系の神経解剖学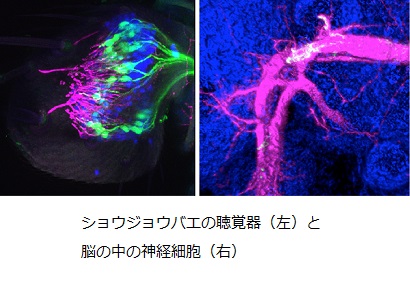 私たち人間を含めた多くの動物は、コミュニケーション手段として音を利用しています。コミュニケーションに使われる音の多くは種に固有のパターンを持つため、その伝達が成立するためには、受け取った音の意味を理解する、といった脳での情報処理が必要です。では、脳の中のどのような神経回路の組み合わせがこのような情報処理を担っているのでしょうか。私たちは、神経回路レベルで脳を理解するための優れたモデル動物であるショウジョウバエを利用して、音情報処理を担う聴覚神経細胞の包括的な同定と神経回路地図の作製を行っています。
■ 音のリズムを聞き分ける神経メカニズムの研究音のリズムを聞き分けることは、ヒトを含むさまざまな動物のコミュニケーションにおいて必須の能力です。ショウジョウバエは、ヒトとは全く異なる外見をしていますが、ヒトと同様に音のリズムの僅かな違いを聞き分けることができ、またヒトと共通した脳内メカニズムを備えていることがわかっています。私たちのこれまでの研究で、ショウジョウバエの脳内のニューロンが、他のニューロンの応答を絶妙に調節する「ブレーキ」の役割を果たすことで、音に対する過剰な反応が起こらないように制御していることがわかりました。この脳内メカニズムをさらに解析することで、ヒトの会話や音声認識にも共通する「音のリズムを分析する脳内メカニズム」の解明に繋げたいと考えています。
■ 音を識別する能力を後天的に獲得する学習機構鳥の歌学習や人間の言語学習は、音パターンを識別する先天的な脳内機構と幼少期での聴覚経験で成熟する後天的な脳内機構とが協調して働くことで実現されます。 私たちは近年、ショウジョウバエも同様のメカニズムで正しい歌を学習することを発見しました。 研究対象としてのショウジョウバエは、研究データを得るスピードの速さや遺伝子操作技術の豊富さを特徴とし、動物一般に共通する神経メカニズムを多く備えています。 私たちはこの利点を生かして、歌や言語学習を担う神経機構や分子機構を解明するための最も単純なモデル系として、多彩な実験操作が可能なショウジョウバエを用いる、という新たな研究戦略を展開しています。
■ 蚊の配偶行動を制御する神経機構の解明と応用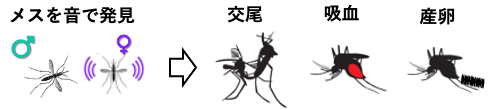 蚊は吸血することで感染症を媒介し、毎年数十万人規模で人を死に至らしめます。蚊の吸血は、交尾後のメスが産卵のための栄養源を得る行動です。よって、蚊の被害を防ぐ方策の一つとして、配偶行動への介入が考えられます。そこで私たちは、日本を含むアジア地域においてデング熱やジカ熱などを媒介するヒトスジシマカや、熱帯地方でそれらの感染症を媒介するネッタイシマカなどの蚊を対象にして、その配偶行動を成立させる神経基盤の解明に挑んでいます。特に、配偶行動に重要な役割を果たすと予想される聴覚系に着目して、その機能や特性を行動や神経細胞のレベルで解明することを目指しています。蚊の配偶行動を聴覚を中心に据えて理解することで、新たな蚊の繁殖制御法の開発に繋がる可能性があります。 ■ HFSP2021 採択課題: 蚊の聴覚コミュニケーションの解読 Decoding acoustic communication in mosquitoes : From distortion products to vector control
■ 行動の進化をもたらす遺伝子と神経回路の変化を特定する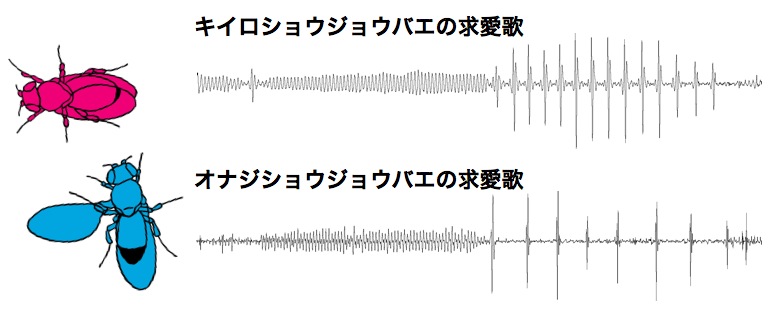 動物の行動は可塑的であると同時に、ある種や系統群を特徴づけるのに非常に有用な形質のひとつでもあります。進化の過程でしばしば行動変化が形態変化に先んじるように見えることから、行動は、形態と相互作用しながら、ときには形態を先導して、進化してきたと考えられています。近年、進化発生学Evo-devoの発展により、形態進化の分子機構については徐々に理解が深まってきましたが、一方で「新たな行動がいかにして獲得されるのか」や「行動の種特異性はどのように成り立っているのか」に関しては、ほとんど理解が進んでいません。
研究業績へ
Copyright (C) Kamikouchi laboratory All rights reserved.
|
 ショウジョウバエは求愛する時に、「求愛歌」と呼ばれる種に固有の羽音を奏でます。このような音の情報処理を担う脳の神経機構の理解を目指して、私たちは、ショウジョウバエの聴覚神経回路の網羅的な同定解析を進めています。今までの私たちの研究から、ショウジョウバエの聴覚器から脳へ投射する一次神経細胞が形成する神経回路地図が世界で初めて明らかになりました。また、二次神経細胞が形成する神経回路の構造を調べてみると、私たち哺乳類の聴覚系と類似性を持つことも分かりました。このことは、ショウジョウバエを用いた聴覚研究が、私たち自身の聴覚システムの理解にも貢献しうることを示しています。現在は、これらの神経細胞から情報を受け取る、より高次な神経細胞の体系的な同定を進めることで、ショウジョウバエ聴覚系を構成する全神経回路の全体像の包括的理解を目指しています。
ショウジョウバエは求愛する時に、「求愛歌」と呼ばれる種に固有の羽音を奏でます。このような音の情報処理を担う脳の神経機構の理解を目指して、私たちは、ショウジョウバエの聴覚神経回路の網羅的な同定解析を進めています。今までの私たちの研究から、ショウジョウバエの聴覚器から脳へ投射する一次神経細胞が形成する神経回路地図が世界で初めて明らかになりました。また、二次神経細胞が形成する神経回路の構造を調べてみると、私たち哺乳類の聴覚系と類似性を持つことも分かりました。このことは、ショウジョウバエを用いた聴覚研究が、私たち自身の聴覚システムの理解にも貢献しうることを示しています。現在は、これらの神経細胞から情報を受け取る、より高次な神経細胞の体系的な同定を進めることで、ショウジョウバエ聴覚系を構成する全神経回路の全体像の包括的理解を目指しています。