Bannai et al. (2009) Neuron
62(5):670-82; Niwa et al. (2012) PLoS
ONE 7(4) e36148
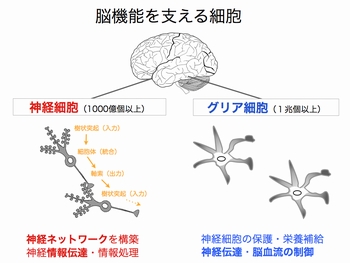
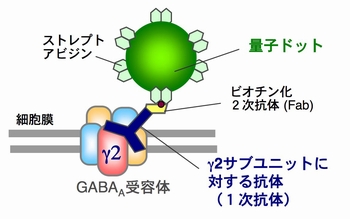
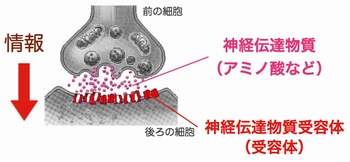
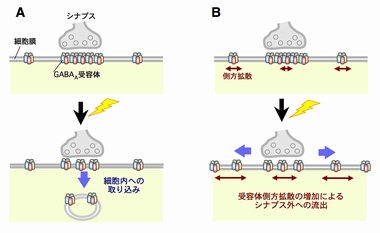
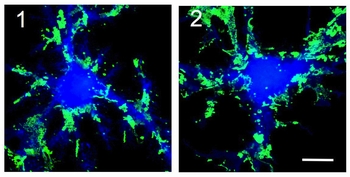
神経細胞と神経細胞は「シナプス」という特殊な構造(図3)を介して,情報を伝達します.このシナプスにおける情報伝達がいろいろな刺激によって強化されたり,弱まったりすることが,記憶・学習の基礎になっています.シナプスの情報伝達の強さが変化する仕組みを明らかにするために,我々は情報を前の細胞から受け取る役割のタンパク質「神経伝達物質受容体(図3,赤)の一種,GABAA受容体1分子を量子ドットで標識しその動きを顕微鏡で追跡しました.すると,シナプス伝達が弱くなる条件では,GABAA受容体の細胞膜上で動きが著しく増加していることが判明しました(動画2).一方,細胞膜上に存在する受容体の量は変わっていませんでした.すなわち,GABAA受容体が細胞内に取り込まれたり壊されたりするのではなく,細胞膜上でGABAA受容体の動きが増してシナプスから出て行きやすくなるため,結果的にシナプスの受容体の数が減りシナプス伝達が弱くなることがわかりました(図4).この研究結果は,シナプス伝達効率を「細胞膜上での受容体の動き」という物理現象が決めていることを意味しています.(Bannai
et al. Neuron 2009)
また,これまGABAARでGABAARの側方拡散制御に関わると信じられて来た唯一の足場タンパク質gephyrinが,シナプス可塑性時のGABAAR側方拡散変化の過程に無関係であるという意外な事実も発見しました(Niwa
et al. PLoS ONE 2012)
2. Niwa F, Bannai H#, Arizono
M, Fukatsu K, Triller A#, Mikoshiba K# “Gephyrin-independent
mobility
and clustering during plasticity.” PLoS ONE
7: e36148. (2012)
3. Tamamushi S, Nakamura
T, Inoue T, Ebisui E, Sugiura K, Bannai H, Mikoshiba K#.
Type 2
inositol 1,4,5-trisphosphate receptor is predominantly
involved in agonist-induced Ca(2+) signaling
in Bergmann glia.Neurosci
Res. 74: 32-41. (2012)
本論文の内容は表紙に採用された.
4. Nakamura H, Bannai H,
Inoue T, Michikawa T#, Sano M, Mikoshiba K#. “Cooperative
and stochastic calcium releases from multiple calcium
puff sites generate calcium microdomains in intact HeLa
cells.” J Biol Chem
287: 24563-24572. (2012)
5. Renner M, Schweizer C,
Bannai H, Triller A, Levi S#. “Diffusion barriers constrain
receptors at
synapses.” PLoS ONE 7: e43032.
6. Fukatsu K, Bannai H, InoueT,
Mikoshiba K#. ”Lateral diffusion of inositol 1,4,5-trisphosphate
receptor type 1 in Purkinje cells is regulated by calcium
and actin filaments.”J
Neurochem, 11:
1720-1733. (2010)
7. Bannai H, Levi S, Schweizer
C, Inoue T, Launey T. Racine V, Sibarita J.B, Mikoshiba
K,Triller A#. “Activity-dependent tuning of inhibitory
neurotransmission based on GABAAR diffusion
dynamics.” Neuron
62:670-682. (2009) 本論文の内容は表紙に採用された.
8. Levi S, Schweizer C, Bannai
H, Pascual O, Charrier C, Triller A#. “Homeostatic regulation
of synaptic GlyR numbers and lateral diffusion.”Neuron
59:261-273. (2008)
9. Bannai H, Levi S, Schweizer
C, Dahan M, Triller A#. “Imaging the lateral diffusion
of membrane molecules with quantum dots.” Nature
Protocols 1:2628-2634. (2006)
10. Fukatsu K, Bannai H,
Inoue T#, Mikoshiba K. “4.1N binding regions of inositol
1,4,5-trisphosphate receptor type 1.”Biochem
Biophys Res Commun 342:573-576. (2006)
11. Tateishi Y, Hattori M#,
Nakayama T, Iwai M, Bannai H, Nakamura T, Michikawa T,
Inoue T, Mikoshiba K. “Cluster formation of inositol 1,4,5-trisphosphate
receptor requires its transition to open state.” J
Biol Chem 280:6816-6822. (2005)
12. Bannai H, Fukatsu K,
Mizutani A, Natsume T, Iemura SI, Ikegami T, Inoue T#,
Mikoshiba K. “An RNA-interacting Protein, SYNCRIP (Heterogeneous
Nuclear Ribonuclear Protein Q1/NSAP1) Is a Component of
mRNA Granule Transported with Inositol 1,4,5- Trisphosphate
Receptor Type 1 mRNA in Neuronal Dendrites.” J
Biol Chem 279:53427-53434. (2004)
Laboratory of Brain Function and Structure;
Division of Particle and Astrophysical Science, Graduate School of
Science of Nagoya University
Copyright (c) 2011 Nagoya University . All Rights Reserved.