PROJECTS
新しい分泌型ペプチドホルモン−受容体ペアの探索と下流シグナル伝達経路の解析
私たちは,ゲノム情報から短鎖の分泌型ペプチドホルモンをコードしうる遺伝子群をin silicoで絞り込み,その成熟型ペプチド構造を質量分析により決定し,さらに受容体候補発現ライブラリーを用いた結合実験から受容体を同定するという戦略で,新規ペプチドホルモン—受容体ペアの探索を進めてきました.限定分解(プロセシング)を経て一部の配列だけが切り出され,最終的な成熟型ペプチドとなるタイプのペプチドホルモンの場合,分子進化の選択圧がかかる成熟型ペプチド部分は高度に配列保存され,機能に関係しない部分には多くのアミノ酸置換や欠失が蓄積するという経験則があります.これを目印とした探索により,根端分裂組織のパターニングに関わるRGF(Science 2010),一部の根の窒素欠乏を他の根からの吸収で補うための全身的窒素要求シグナリングに関わるCEP(Science 2014),根におけるイオンや水の拡散障壁として機能するカスパリー線の形成に関わるCIF(Science 2017),成長とストレス応答のトレードオフを制御するPSY(Science 2022)など,植物の成長や環境応答に関わるホルモン群を発見してきました.
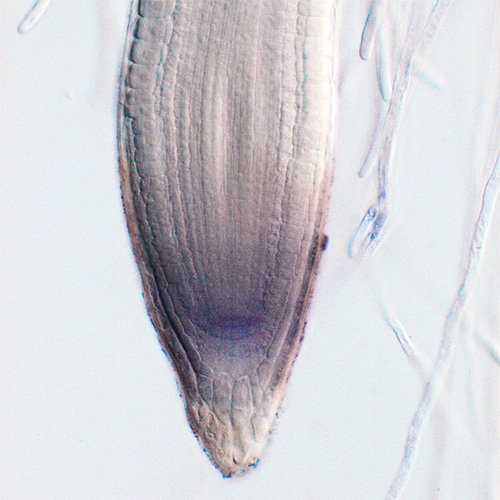
これらのペプチドホルモンのうち,RGF,CIF,PSYなどはチロシン硫酸化という翻訳後修飾を受けたペプチドで,その生理機能解明の過程では,チロシン硫酸化酵素の欠損株(PNAS 2009)が大きな役割を果たしました.現在では,これらのペプチドホルモン群が,受容体に認識された後に,どのようなシグナル伝達経路でターゲット遺伝子群の発現を誘導するのかを解析しています.
篩管移行性の非分泌型ペプチドシグナルの探索と機能解明
植物には,根から吸収した栄養分を地上部に運ぶ通り道である道管と,葉から光合成産物などを頂芽や根に送る篩管がありますが,両者は長距離情報伝達の場としても重要であることが近年明らかになってきました.特に興味深いのは,篩管を通って器官間を長距離移行するペプチド群です.
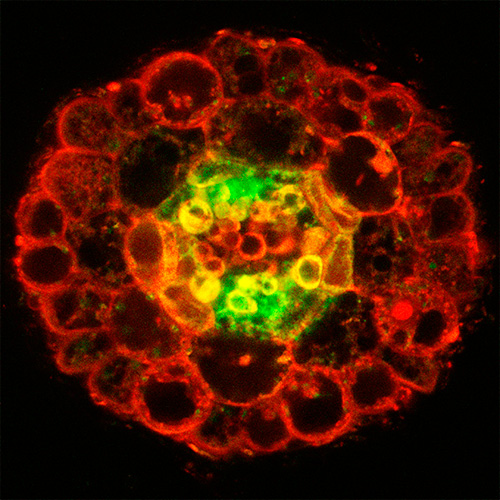
最近の私たちの研究により,葉でつくられるCEPDやCEPDL2などの非分泌型ペプチドが,篩管を通って根へ移行し,硝酸取り込み輸送体の発現を調節していることが明らかとなっています(Nature Plants 2017, Nature Commun. 2020).篩管は生きた細胞が連なってできた通道組織で内部は原形質で満たされていますが,篩管の細胞と細胞の境界には篩孔と呼ばれる穴が多数存在するため,内部の原形質は細胞間を移行できます.そのため,篩管内の非分泌型ペプチドは器官間を長距離移行できるのです.これは動物にはない植物独自のしくみで,まだ分からないことが数多く残されています.
私たちは,類似の移行能を示すような非分泌型ペプチド群に着目すれば,植物の地上部と地下部のダイナミックな情報のやりとりが明らかになる可能性があると考えています.そこで,特殊な方法で篩管を含む維管束組織を葉から単離し,トランスクリプトームを行なって篩部に発現する非分泌型ペプチド群を絞り込み,個々の器官間移行性や生理機能を解析しています.また,細胞内を移行するこれらの非分泌型ペプチドは細胞膜上の受容体に認識されるとは考えにくいので,器官間を移行した後にどのようにターゲット細胞で特定の遺伝子発現を誘導するのかについても研究しています.
糖転移酵素に着目したグリコバイオロジー
ヒドロキシプロリン(Hyp)にアラビノース糖鎖が付加するHypアラビノシル化は,一部のペプチドホルモンや細胞外タンパク質の機能に重要な役割を果たす翻訳後修飾です.私たちは,この修飾反応に関わる糖転移酵素(Hydroxyproline O-arabinosyltransferase: HPAT)を,シロイヌナズナの細胞の膜タンパク質画分から,精製・同定することに成功しています(Nature Chem. Biol. 2013).HPATを欠損させた植物体では,細胞壁厚の顕著な減少,花成の促進,および花粉管伸長異常による不稔など,栄養成長および生殖成長の両方に様々な異常が観察されています.これらの結果は,どこでどのようなアラビノシル化ペプチドやタンパク質が機能しているかを知る上で重要な情報となり,新しい研究の出発点になります.
実際,マメ科植物におけるHPATの変異体では,窒素固定に関わる根粒の数が増加することが知られており,これは根粒の数を調節するペプチドホルモンのアラビノース糖鎖が失われることによる機能欠損が原因であることが明らかになりました(Nature Commun. 2013).トマトでもHPATの変異体では,メリステム幹細胞の異常な蓄積による花や果実の数の増加が観察されており,これもメリステムの幹細胞数を制御するペプチドホルモンのアラビノース糖鎖が失われることによる機能欠損が原因であることが確かめられています.
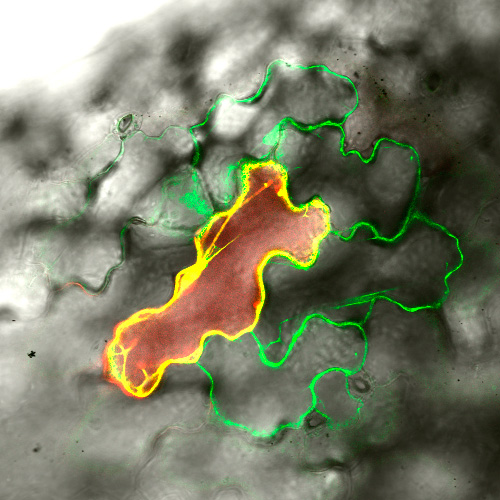
また,私たちは植物で大きなファミリーを形成しているアラビノガラクタンプロテイン(AGP)の糖鎖付加の第1段階を担う糖転移酵素(Hydroxyproline O-galactosyltransferase: HPGT)の精製・同定にも成功しています(Plant J. 2015).AGPは,植物の成長や分化に重要な役割を果たすと考えられているプロテオグリカンですが,遺伝子重複のために解析が難しいことでも知られています.HPGTの欠損株では,すべてのAGPへの糖鎖付加ができなくなるため,遺伝子重複の障壁を乗り越えてAGP分子群の機能の全体像に迫ることができる可能性があります.私たちはこれまでに,植物体の矮化,側根や根毛の伸長促進,根端細胞の肥大,稔性の低下,原形質連絡の異常によるシンプラスト輸送の昂進などの表現型を観察しています(Plant J. 2015, 2023).これらの結果を手がかりにして,AGPの機能解析を進めていきたいと考えています.
タンパク質脱リン酸化酵素の基質探索
私たちは,篩管移行性の非分泌型ペプチドシグナルのひとつであるCEPDL2の機能解析の過程で,根に移行したCEPDL2が機能未知のタンパク質脱リン酸化酵素(後にCEPHと命名)の発現を誘導することを見出しました.このCEPHの機能を明らかにするには,その基質を同定する必要があります.
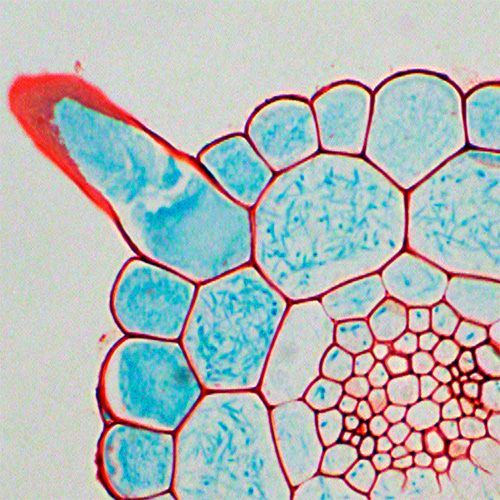
そこで,独自に改良した15N代謝ラベル定量リン酸化プロテオミクス系を用いて,CEPHの欠損株中でリン酸化レベルが上昇するタンパク質を探索しました.その結果,CEPHは根における主要な硝酸イオン輸送体であるNRT2.1の501番目のSer残基を脱リン酸化しており,これにより硝酸イオンの取り込み活性がONになることが明らかになりました(Nature Plants 2021).
タンパク質脱リン酸化酵素は,様々なシグナリング系で鍵となる役割を担う例が知られている一方で,機能未知のものがまだ多数残されています.特に,CEPHが属するPP2Cファミリーのタンパク質脱リン酸化酵素はシロイヌナズナに80個存在しており,ホルモン応答や環境応答に関わることが明らかにされたものも含まれていますが,全体の70%はまだ基質が明らかになっていません.2倍差以下の変動でも検出できる15N代謝ラベル定量リン酸化プロテオミクスで,それぞれの基質を同定できれば,植物成長に関わる新しいシグナル経路やしくみを発見できる可能性があります.