論文紹介
| 研究代表者 |
伊藤純一
佐藤 豊 |
所 属 |
東京大学大学院農学生命科学研究科
名古屋大学大学院生命農学研究科 |
| 著 者 |
Jun-Ichi Itoh, Ken-Ichiro Hibara, Yutaka Sato, and Yasuo Nagato
(伊藤純一、桧原健一郎、佐藤豊、長戸康郎) |
| 論文題目 |
Developmental Role and Auxin Responsiveness of Class III Homeodomain Leucine Zipper Gene Family Members in Rice
(イネのクラスIIIホメオドメインロイシンジッパー遺伝子ファミリーの発生における役割とオーキシンの反応性) |
| 発表誌 |
Plant Physiology. 147:1960-1975, 2008 |
| 要 旨 |
シロイヌナズナにおいてクラスIIIホメオドメインロイシンジッパー遺伝子(HD-ZIP III)は発生過程において極めて重要な役割を果たしているが、イネ(単子葉植物)における機能は明らかではない。そこで、イネにおける5つのHD-ZIP III 遺伝子の発生過程における発現パターン、過剰発現体の表現型、オーキシンによる反応性を解析し、イネにおけるHD-ZIP III の発生における役割について考察を行なった。in situ hybridizationによる発現解析では、5つのHD-ZIP IIIうち4つの遺伝子は茎頂分裂組織(SAM)、葉原基の向軸側、維管束の木部の細胞などで発現が観察された。過剰発現体の表現型観察では、本来向軸側にできる組織が背軸側に分化した葉や、極性を失った糸状の葉原基などが観察された。これらのことから、イネのHD-ZIP III遺伝子はシロイヌナズナと同様に、SAMの形成や側生器官における向軸側のidentityの決定に関与していることが示唆された。一方で、過剰発現体において、葉縁に特徴的な膜状の構造や糸状の葉原基が、本来の葉原基やSAMとは独立に形成されたことから、イネのHD-ZIP III遺伝子は葉縁の形成や葉原基分化にも機能を持っていることが推測された。
続いて、HD-ZIP III遺伝子のオーキシン反応性について調査した。植物体全体においてはいずれのHD-ZIP III遺伝子も顕著なオーキシン反応性を示さなかったが、in situ hybridizationの観察から、SAMにおいてのみ強いオーキシン誘導性が見られた。このことからHD-ZIP III遺伝子はSAMにおいて、オーキシンに依存した葉原基分化過程に何らかの働きを持っていることが予想された。
|
|
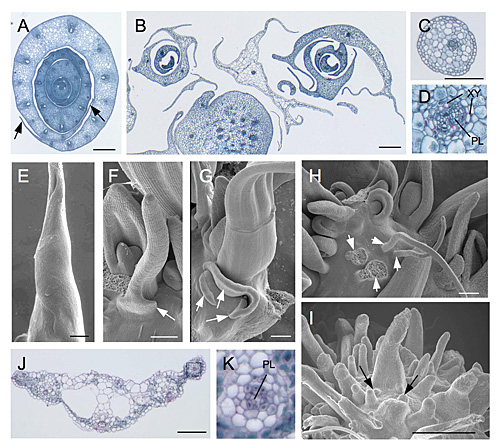
図1 miR166耐性OSHB遺伝子の過剰発現体の表現型。
A, E. 野生型、B, C, D, F, G, H, I. 変異型OSHB3遺伝子の過剰発現体。J, K. 変異型OSHB5遺伝子の過剰発現体。B. 多数の膜状の構造(野生型の葉縁に対応)が認められる。C. 糸状になった葉の断面。D. 維管束の極性も異常となっている(XY:木部、PL:師部)。E. 野生型葉原基の電子顕微鏡写真。F, G. 膜状の構造(矢印)が認められる。H, I. 糸状の葉原基(矢印)が認められるがSAMが認められない。J, K. 葉の外形は比較的正常だが、維管束の極性に異常が認められる。 |
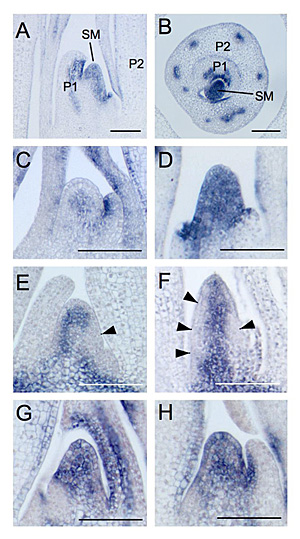 図2 オーキシン処理による OSHB遺伝子の発現。
A, B, D. 1週間の2,4-D処理後のSAMにおける OSHB3遺伝子の発現。SAM全体で発現が認められる。(SM: SAM, P1, P2: 葉原基)C. 無処理のSAMにおける OSHB3遺伝子の発現。SAMの中心のみで発現が見られる。E. 野生型における OSH1の発現。SAMの葉原基分化予定領域(矢頭)で発現抑制が観察される。F. 1週間の2,4-D処理後のSAMにおける OSH1の発現。SAMの広い範囲(矢頭)で発現抑制が観察される。G, H. 6時間の2,4-D処理後のSAMにおける OSHB3遺伝子の発現。SAM全体で発現が認められる。 |
|
| 研究室HP |
|
|