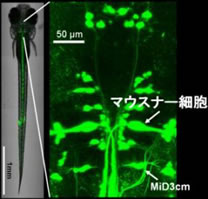
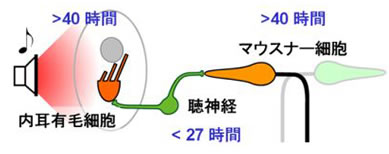
M細胞の重要性が強調されていながら,M細胞を破壊されたサカナでも逃避運動ができるというパラドックスがあることが今から30年近く前から分かっていました.やがて,この逃避運動はM細胞以外のRSニューロン(図2)が引き起こすと考えられるようになりましたが,そもそも逃避運動中のRSニューロンの活動は,M細胞以外に計測された例がありませんでした.古典的な電気生理学的手法では,運動中に活動したニューロンを同定することなどほとんど不可能だったため(M細胞の活動が同定できるのは,ごく例外的なのです).そこで私たちは,RSニューロンの活動と逃避運動との相関を調べるため,逃避運動中のゼブラフィッシュからRSニューロン活動をin vivoでカルシウムイメージングするシステム(図4)を用いて,この問題を克服ました.
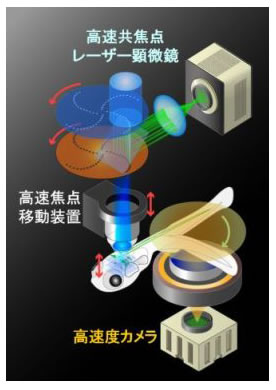
このシステムは,以前私たちが開発して,ゼブラフィッシュ生体内での抑制性回路のはたらきをカルシウムイメージングした際(当時,これは世界初の試みでした)に用いた共焦点レーザー顕微鏡(Takahashi, Narushima and Oda, J. Neurosci., 2002)の下に,新しい光学系と高速度カメラを取り付けたものです.これによって,頭だけ寒天の中に拘束した稚魚の脳内のRSニューロンを,共焦点レーザー顕微鏡で光学計測しながら,同時に起こった,自由に動く尾の逃避運動を計測することができるようになりました.このときRSニューロンにはカルシウム感受性の蛍光色素を取り込ませてあるため,ニューロン活動に対応したカルシウム濃度変化を蛍光強度の変化としてイメージングできます (原理図).もうひとつ,世界初の工夫があります.ニューロンのイメージングと同期して焦点面を高速で移動させることで,深さが異なる複数のニューロンでも,活動を同時に計測できるようにしたのです.
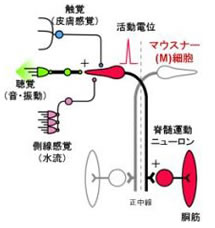
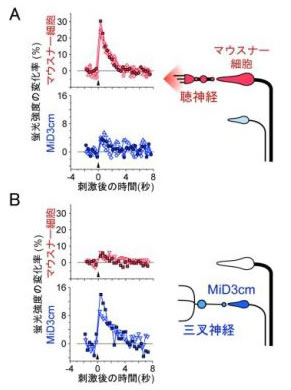
このような光学系を使って逃避運動中のM細胞や他のRSニューロンの活動を調べた結果,私たちはゼブラフィッシュには異なる感覚入力で駆動される,2種類の逃避運動回路があることを見出しました.すなわち,聴覚刺激はM細胞を1回発火させて最も素早く逃避運動を開始する(データの一例)一方,頭への触刺激は三叉神経を活動させて逃避運動を開始するのですが,このときM細胞は発火せず,かわりにM細胞と形態の似たMiD3cmというRSニューロンが大きな活動を示すことが分かりました(図5)
この成果は,J. Neurosci. (Kohashi and Oda, 2008)に発表しました.この論文は,逃避運動中の非M細胞の活動,とくに非M型逃避の実体をはじめて明らかにした点が評価されて,論文掲載号で注目論文として取り上げられたり,J. Exp. Biol.で紹介されました.国内でも,朝日,中日,日刊工業新聞で報道された他,NHKニュースや教育テレビでも取り上げられました.. |