生物が進化の過程で脳(集中神経系)を獲得したときから「脳は分節(セグメント)構造をなしている」。最も原始的なホヤ幼生の単体節神経系からナメクジウオの多体節神経系になったときに起こった基本構造の重複と変異こそが、脳の進化の出発点であり、このプロセスは哺乳動物まで保存される。脳が分節構造をとったことが、脳の機能原理にも重要な方向を与えたのではないだろうか?
私達は脳の分節構造に注目して、分節構造に基づいたニューロンの機能分化とそれらが相互に結合してつくる機能回路を調べている。研究対象はゼブラフィッシュやキンギョの逃避運動をコントロールする後脳の網様体脊髄路ニューロン群である。
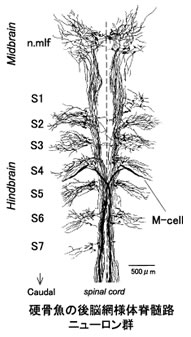
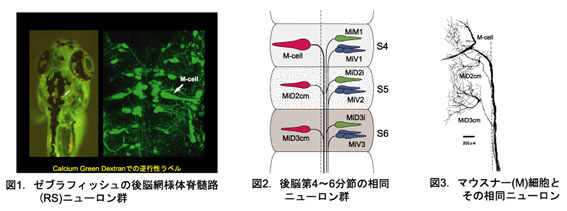
分節構造は発生期には脳のくびれとして明確に見分けられるが、通常成熟とともに不明瞭になる。ところが硬骨魚の後脳網様体脊髄路(RS)ニューロン群は、成魚においても明確な分節構造を保っている。ゼブラフィッシュやキンギョの後脳には20~30種のRSニューロンが総数100~200個存在し、脳の前後軸方向に7つの分節を形成する。さらに、形態学的によく似た相同RSニューロンが隣接する分節に存在するという興味深い配列を示す。最大の大きさをもつマウスナー(M)細胞が魚の逃避運動をトリガーすることが古くから知られていた。最近、ニューロン活動のイメージングによって、M細胞と形態学的に相同なRSニューロンも逃避運動の発現と制御に関与する可能性が示された。
われわれは、M細胞と相同RSニューロンが互いにどのような電気生理学的特性を持つか、どのような入力投射を受けるか、相互にどのような結合をつくっているか、つくられた後脳RSニューロン回路は魚の逃避運動の制御にどのような役割を果たすかを調べて、脳が分節をつくって発達したことが脳の機能獲得にどのような役割を果たしたかを理解しようとしている。
脳の分節構造に関しては、分節に対応して発現する遺伝子に関して盛んに研究されてきた。後脳ではHoxやKrox20などの転写因子や細胞間相互作用に働くレセプター・チロシンキナーゼなどである。これらの遺伝子相互の関係やマクロスコピックな分節の形成に及ぼす効果は示されているが、個々のニューロンの性質が分節によってどのように決められるかについて明確な答えは得られていない。 われわれは、分節に埋め込まれた個々のニューロンの性質と相互結合が明らかにされれば、分節を決める遺伝子から脳ニューロンの性質と回路の機能までを系統的に調べられると期待している。
脊椎動物の脳は発生初期に前脳・中脳・後脳(菱脳)に領域化され、さらに各領域がセグメント(分節)を形成する。 分節構造は脳の基本構造のひとつである。 脳が普遍的に分節構造をとることは、脳が機能を発揮するうえでどのように反映されているのだろうか? 本研究では、脳の分節構造に基づいたニューロンの機能分化と相互の機能結合を、魚の逃避運動を制御する後脳の神経回路に見出した。
後脳分節に繰り返される形態学的に相同なニューロンの性質と回路構成明らかにする。
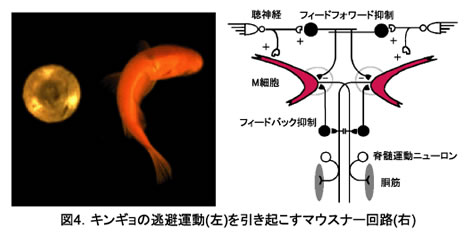
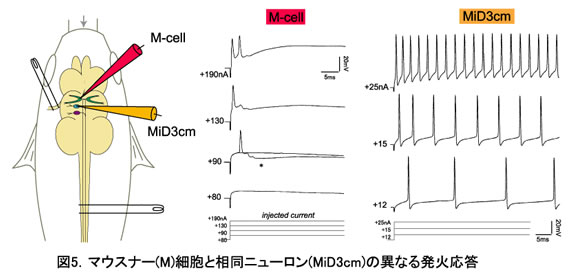
硬骨魚の後脳に存在するRSニューロン(図1)はすべて形態学的に同定され(約30種)、形態学的に相同性の高いニューロンが隣接した分節に繰り返されている(図2)。第4分節には魚の逃避運動(図4)をトリガーし、最大の大きさをもつマウスナー(M)細胞が存在する。 M細胞とその相同RSニューロンの電気生理学的特性、相互の結合および入出力関係を調べ、分節に従った回路構築を明らかにする。
M細胞は強い脱分極電流を通電してはじめて発火し、しかも開始時に1-2発の活動電位を発生するのみである。 一方、その他のRSニューロンは脱分極の大きさに応じたバースト発火を示す。 すなわち、M細胞は入力に対しall-or-nothingに応答し、それ以外のRSニューロンは入力量を反映した発火頻度を出力する。 M細胞の特異的な興奮性は、M細胞の局所抑制回路と膜のカリウムチャネルによって制御されていると考えている。
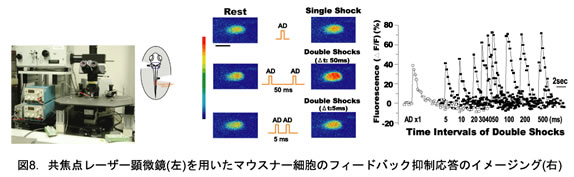
ゼブラフィッシュ稚魚のM細胞にカルシウム指示薬(カルシウムグリーン・デキストラン)を取り込ませ、共焦点レーザー顕微鏡を用いてニューロン活動を高速イメージングした。これまでイメージングでは興奮性応答しか観察されていなかったが、本研究によって抑制性シナプス応答を生体内で光学計測することに初めて成功した。その結果、成熟型逃避運動が発現する受精後4日で既にM細胞のフィードバック抑制性回路が機能していることが明らかにされた。
硬骨魚(キンギョ,ゼブラフィッシュ)の後脳網様体脊髄路(RS)ニューロン群は、脳の吻尾軸方向に7つの分節をつくって並んでいる。第4分節には魚の逃避運動をトリガーする巨大なマウスナー(M)細胞が存在し、その形態学的相同ニューロンは隣接する第5~6分節に繰り返される。
1.In vivo細胞内記録(キンギョ)から、M細胞と相同RSニューロンの間には膜の興奮性が大きく異なることが示された。逃避運動の開始と左右方向を決定するM細胞が単発の活動電位を発生するには、M細胞のフィードバック抑制性回路に加えて、特異的な電位依存性カリウムチャネル発現が重要な役割を果たすと考えられる。
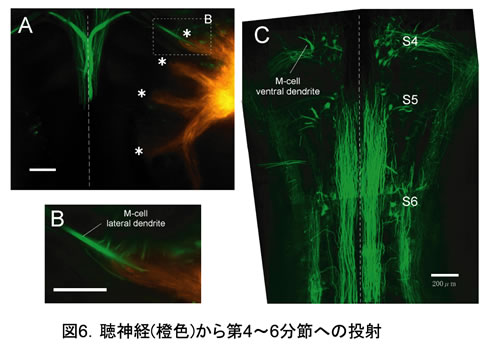
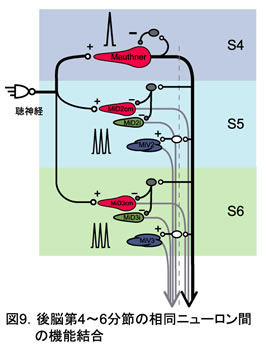
2.逃避運動の誘導に必要な聴神経は、M細胞と相同RSニューロンの存在する第4~6分節に限局して投射していた。
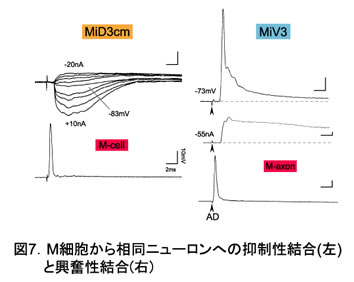
3.M細胞から第5~6分節の相同RSニューロンへの一方向性の結合が明らかになった。しかも、M細胞と相同性の高い背側ニューロン(MiD)へは抑制性投射、相同性の低い腹側ニューロン(MiV)へは興奮性投射という。形態学的相同性を反映した結合が見出された(図9)。
4.M細胞の活動を強く制御する局所抑制性回路の働きを、ゼブラフィッシュ稚魚からin vivoイメージングすることに成功し、成熟型逃避運動が発現し始める受精後4日で既にフィードバック抑制が機能していることを見出した。
分節に重複した相同ニューロンから形成された神経回路網が、M細胞だけでは達成されない俊敏で多様な逃避運動パターンを発現すると考えられる。